














Рис. 1. Причудливый облик ресничных личинок. Верхний ряд — личинки типа трохофоры, нижний — типа диплеврулы. Внутрипорошицевые — небольшая группа морских или пресноводных сидячих первичноротых (раньше их считали мшанками). Остальные пояснения — в тексте. Иллюстрации из статьи: C. Nielsen, 1985. Animal phylogeny in the light of the trochaea theory
Трохофоры и диплеврулы — морские личинки первичноротых (кольчатые черви, моллюски) и вторичноротых (иглокожие, полухордовые) животных. Как правило, они совершенно не похожи на свои взрослые формы, да и друг от друга они тоже сильно отличаются. Откуда взялись все эти отличия? И что они означают с эволюционной точки зрения? Эти вопросы интригуют зоологов уже примерно 140 лет. Сейчас предложен эволюционный сценарий, согласно которому у трохофор и диплеврул всё-таки (вопреки некоторым сомнениям) есть общий план строения, восходящий к общему предку всех билатерально-симметричных животных.
Полупрозрачный мир
Одна из самых больших загадок зоологии, беспокоящая исследователей уже не первый век, заключается в следующем. Почему полупрозрачные планктонные личинки морских беспозвоночных, в изобилии плавающие по Мировому океану, так поразительно отличаются от взрослых животных, в которых они потом превращаются? Многие из этих личинок имеют собственные названия: трохофора — личинка кольчатых червей и моллюсков, актинотроха — личинка форонид, торнария — личинка кишечнодышащих, плутеус — личинка морских ежей и офиур, бипиннария — личинка морских звезд, аурикулярия — личинка морских огурцов и так далее. Несведущий человек ни за что не угадает, например, в трохофоре будущего многощетинкового кольчатого червя или в плутеусе — будущего морского ежа (рис. 1). Планктонные личинки очень разнообразны, однако их разнообразие вовсе не хаотично: давно известно, например, что иногда невероятно сходными личинками обладают животные, у взрослых стадий которых нет буквально ничего общего (по крайней мере, внешне). Одним словом, морские личинки — это целый мир, имеющий свои загадочные закономерности, на раскрытие которых не жалко потратить жизнь. И действительно, биологи с такими интересами находились и находятся. Но, как известно, «ничто в биологии не имеет смысла кроме как в свете эволюции», поэтому главный вопрос тут: откуда все эти личинки вообще взялись?
Возможных ответов по большому счету есть два. Один из них высказал несколько десятилетий назад петербургский эмбриолог, профессор Петр Павлович Иванов: «Личинка появляется в развитии животного потому, что питательного материала, накапливающегося в ооците в период желткообразования, недостаточно для полного завершения зародышем своего развития, и он выходит из яйца на более или менее ранней эмбриональной стадии» (П. П. Иванов, 1937. Общая и сравнительная эмбриология). По этой версии, древнейшие многоклеточные животные ползали по дну, а никакой личинки у них вообще не было. Иными словами, развитие было прямым: из яйца вылуплялась миниатюрная версия взрослого животного. И только потом вылупление сдвинулось на гораздо более ранние стадии — то есть, попросту говоря, вылупляться стал эмбрион. Вот почему личинки выглядят так неожиданно. «Чем более ранней является стадия, на которой происходит выход из оболочки, тем меньше признаков, сходных со взрослой формой, имеет личинка», — пишет Иванов. Например, микроскопические личинки иглокожих и многих кольчатых червей вылупляются очень рано, поэтому неудивительно, что они не напоминают взрослых особей вообще ничем.
Склонность к возникновению личинок могла быть связана и с проблемой расселения. Для морского животного, ползающего по дну, а тем более сидячего, планктонная личинка — это единственный шанс расселиться на большое расстояние. Зато эффективность такого расселения огромна, ведь океанские течения могут переносить личинок на тысячи километров. Поскольку желтка (то есть запасных питательных веществ) для поддержания жизни длительно плавающей личинки, как и говорил Иванов, не хватало, личинка была вынуждена научиться кормиться самостоятельно. Из лецитотрофной — питающейся желтком — она стала планктотрофной, питающейся более мелкими планктонными организмами. Австрийские зоологи Герхард Хашпрунар (Gerhard Haszprunar), Луитфрид Сальвини-Плавен (Luitfried Salvini-Plawen) и Райнхард Ригер (Reinhard Rieger), основательно изучившие эту проблему, были уверены, что планктотрофные личинки независимо возникали в разных эволюционных ветвях животных, причем много раз (G. Haszprunar et al., 1995. Larval planktotrophy — a primitive trait in the Bilateria?). Сальвини-Плавен даже назвал широко распространенную тенденцию к образованию таких личинок особым термином — «нормативный параллелизм». Впрочем, у многих беспозвоночных личинки и по сей день остаются лецитотрофными, то есть, попросту говоря, непитающимися.
Альтернативная идея состоит в том, что жизненный цикл многоклеточных животных с самого начала был двухфазным: планктотрофная личинка, живущая в толще воды, и донная взрослая форма. Эту идею отстаивал, например, известный шведский зоолог Госта Егерстен (Karl Gustav «Gosta» Magnus Jagersten). Он считал, что все переходы к непитающейся лецитотрофной личинке или к безличиночному прямому развитию произошли гораздо позже. Если эта версия верна, то надо ожидать, что личинки современных морских беспозвоночных сохраняют очень древние признаки, возможно, унаследованные еще от общего предка всех животных.
Главный из таких признаков очевиден: это плавание с помощью ресничек. Ресничное движение — чрезвычайно древнее, оно унаследовано прямо от одноклеточных, в отличие от пришедшего ему на смену мышечного движения, которое по определению возможно только для многоклеточного тела. У взрослых современных животных ресничное движение сохраняется не так уж часто. Примеры групп, в которых оно до сих пор есть, — гребневики, ресничные черви, коловратки. Но для древнейших многоклеточных животных, у которых мышцы еще не появились, ресничное движение, скорее всего, было единственно возможным вариантом. Современных ресничных личинок иногда называют первичными. Предполагается, что в некоторых группах первичная личинка была потеряна и возникла форма с прямым развитием, которая потом по каким-то причинам «изобрела» личиночную стадию заново. Такая личинка называется вторичной. Типичный пример вторичной личинки — науплиус, планктонная личинка ракообразных, не имеющая никаких ресничек и плавающая с помощью конечностей.
Ясно, что «ивановская» и «егерстеновская» версии противоречат друг другу. С «ивановской» точки зрения деление личинок на первичных и вторичных бессмысленно: они все — вторичные. И никакой информации о древних предках личинки в этом случае нести не обязаны. Чтобы понять, кто тут прав, было бы неплохо для начала разобраться в том, как вообще возникли многоклеточные животные.
Вопрос подвижности
Сценарий происхождения многоклеточных животных недавно обсуждался на «Элементах» (см. «Первичность губок» опережает по очкам «первичность гребневиков», «Элементы», 12.11.2019), поэтому сейчас можно сказать о нем кратко. Если отбросить устаревшие мнения, интересные ныне только историкам науки, то мы имеем право с полной уверенностью утверждать, что многоклеточные животные произошли от каких-то колониальных протистов, относившихся к супергруппе заднежгутиковых. Проще говоря, любое многоклеточное животное — это бывшая колония одноклеточных. Однако гипотезы колониального происхождения животных делятся на две группы. Большинство исследователей во все времена было уверено, что предок многоклеточных животных мог быть только активно плавающим (в крайнем случае — активно ползающим) существом, которое специализировалось на поступательном движении и именно в движении находило свою добычу. Это — гипотезы первичной мобильности. К ним относятся теория гастреи Геккеля, теория фагоцителлы Мечникова и некоторые другие, не столь популярные сейчас идеи.
Гипотезам первичной мобильности противостоят гипотезы первичной седентарности, согласно которым общий предок всех животных был прикреплен ко дну. На первый взгляд такое мнение выглядит странно: мы привыкли ассоциировать животную жизнь с подвижностью, и для этого есть основания. Но еще в начале XX века бельгийский зоолог Огюст Ламэр (Auguste Alfred Lucien Lameere) обратил внимание на то, что способ питания, который скорее всего был у самых примитивных животных — подгон ресничками потоков воды со взвешенными в них мелкими пищевыми частицами — гораздо легче осуществим в сидячем состоянии, когда тело зафиксировано. Губки питаются так до сих пор. Почему не предположить, что первые животные были похожи на них? Иное дело, что морским сидячим организмам нужно как-то расселяться, поэтому в их жизненном цикле довольно скоро (по мнению Ламэра) появились свободноплавающие расселительные стадии — личинки, еще не способные питаться.
Современная молекулярная филогенетика после долгих споров, похоже, все-таки подтвердила, что древнейшая группа многоклеточных животных — это именно губки (если не учитывать загадочные ископаемые формы, родственные связи которых пока неизвестны; см. Подтверждена принадлежность дикинсонии к животному царству, «Элементы», 24.09.2018). У губок встречаются планктонные личинки, похожие на бластулу, одну из самых ранних стадий эмбрионального развития многоклеточных животных. Эти личинки — чисто расселительные и лецитотрофные. У современных губок они не имеют рта и не питаются. Все это отлично подтверждает высказанное сто лет назад мнение Огюста Ламэра. Скорее всего, бластулообразные личинки губок никогда и не питались активно, а были «изобретены» только для расселения: предполагать более сложный ход эволюции тут нет никакой нужды.
Что же было дальше? В наше время бластулообразные личинки характерны для известковых и гомосклероморфных губок, причем с помощью электронной микроскопии удалось показать, что по ряду тонких деталей клеточного уровня эти личинки часто бывают гораздо более похожи на настоящих многоклеточных животных, чем на взрослых губок (см. «Первичность губок» опережает по очкам «первичность гребневиков», «Элементы», 12.11.2019). Напрашивается предположение, что одна из таких личинок перестала проходить метаморфоз и научилась размножаться в личиночном состоянии. Размножение личинок — хорошо знакомое биологам явление, которое называется неотенией. Но неотеническая личинка, переставшая превращаться во взрослую губку, должна была, естественно, освоить самостоятельное питание. Ей пришлось научиться поедать своих более мелких соседей по планктону — стать планктотрофом. Так и появился общий предок настоящих многоклеточных животных (то есть всех животных, кроме губок): бластулоподобный планктонный и планктотрофный организм, который Эрнст Геккель в XIX веке назвал бластеей.
Наследники гастреи
Для реконструкции дальнейших событий вполне можно, не изобретая велосипед, воспользоваться готовой рабочей гипотезой, а именно теорией гастреи того же Геккеля (конкурирующая с ней теория фагоцителлы Мечникова на данный момент выглядит менее вероятной). Одно из полушарий бластеи погрузилось внутрь и образовало второй слой клеток, выстилающий внутреннюю полость. Теперь главной задачей ресничек стало загнать пищевые частицы в эту полость, оттуда они уже никуда не могли деться. Образовавшийся двуслойный организм и называется гастреей. Внутренняя полость гастреи стала первичной кишкой, или архентероном, а ведущее в нее отверстие — первичным ртом.
Геккелевская гастрея (если она существовала) имела голопелагический жизненный цикл, целиком проходящий в водной толще. Кроме того, она была радиально-симметричной, то есть не имела спинной и брюшной сторон. Плавала она, конечно же, с помощью ресничек. Радиально-симметричные голопелагические организмы, плавающие с помощью ресничек, существуют и сейчас. Это — гребневики. От геккелевской гастреи они принципиально отличаются в первую очередь тем, что приобрели нервную систему и мышцы. Для «карьеры» планктонных хищников эти приобретения оказались крайне полезными. Тем не менее гребневики — это, вероятно, самые прямые наследники гастреи, какие только возможны в современном мире. Они никогда не имели никакой симметрии, кроме радиальной, и никогда не использовали никакой другой механизм плавания, кроме биения ресничек (точнее, гребных пластинок, склеенных из множества ресничек). Некоторое исключение составляют ползающие гребневики (см. Платиктениды), но они появились эволюционно недавно, и от них пока больше никто не произошел.
Молекулярная филогенетика подтверждает, что гребневики — самая базальная (близкая к основанию древа) эволюционная ветвь настоящих многоклеточных животных. Они единственные, кто сохранил голопелагический жизненный цикл, свойственный гастрее (но не губкам). Другая большая эволюционная ветвь настоящих многоклеточных животных вскоре перешла к пелаго-бентическому жизненному циклу, взрослая стадия которого живет на дне (рис. 2). Эта ветвь тоже приобрела нервную систему и мышцы и разделилась на две группы: стрекающих и билатерально-симметричных. Последних кратко называют билатериями.

Рис. 2. Происхождение гребневиков, стрекающих и билатерий. Иллюстрация из обсуждаемой статьи в «Онтогенезе», с изменениями
От коралла к медузе
Билатерально-симметричные животные имеют четко дифференцированные стороны тела: спину, брюхо, «право» и «лево». Элементы диссимметрии (например, у улитки) или даже вторичной радиальной симметрии (например, у морской звезды) только дополняют их план строения. Всеми признано, что становление билатеральной симметрии было связано с ползанием по дну. В этих условиях брюшная сторона, обращенная к грунту, стала резко отличаться от спинной, и одновременно возник головной конец, на котором сконцентрировались органы чувств: на дне можно найти гораздо больше пищи, чем в водной толще, но только если активно искать. С этого и началась необычайно богатая событиями эволюционная история билатерально-симметричных животных, от плоских червей до человека.
В учебниках обычно пишут, что у стрекающих (кораллов, медуз, гидр и им подобных) симметрия осталась древней, радиальной. В то же время зоологи всегда — по крайней мере, на протяжении последних ста с лишним лет — прекрасно знали, что у коралловых полипов внутренняя (а иногда и внешняя) симметрия на самом деле скорее билатеральная. Ее задает плоскость щелевидной глотки, которая занимает в теле кораллового полипа весьма существенное место (В. М. Шимкевич, 1923. Биологические основы зоологии). От этого факта не отмахнешься: молекулярная филогенетика свидетельствует, что коралловые полипы образуют как минимум самую базальную эволюционную ветвь стрекающих, а как максимум — базальный эволюционный уровень, через который должны были пройти все представители этой группы. Возникает вопрос: не является ли билатеральная симметрия исходным для стрекающих признаком, а ее полное отсутствие у существ вроде пресноводной гидры — как раз вторичным? Эта проблема уже обсуждалась на «Элементах» (см. Двусторонняя симметрия кораллов древнее радиальной, «Элементы», 26.10.2016). Сейчас достаточно сказать, что возможен эволюционный сценарий, согласно которому и стрекающие, и билатерии произошли от донного билатерально-симметричного ползающего предка (см. рис. 2). Билатерии так и остались ползающими и, разумеется, сохранили свою билатеральную симметрию. А вот предок стрекающих после короткого периода ползающей жизни прикрепился к субстрату. В результате у него произошла вторичная радиализация: прикрепленное хищное животное должно быть готово схватить добычу, с какой бы стороны она ни приблизилась. Так и возникли коралловые полипы.
Упомянутый выше Госта Егерстен считал, что переход к донному образу жизни совершила не гастрея, а бластея. Но тогда элементы билатеральной симметрии должны были бы найтись и у гребневиков, чему нет серьезных подтверждений. У кораллов же они есть точно, причем у всех.
Почти все коралловые полипы — существа или строго сидячие, или малоподвижные. А расселяться им как-то надо. Поэтому в их жизненном цикле сохранилась планктонная стадия — теперь она стала личинкой. У современных одиночных шестилучевых кораллов встречается планктотрофная личинка, очень похожая на гипотетическую геккелевскую гастрею, которая в конце концов оседает на дно (см., например: J. F. Gemmill, 1920. The development of the sea-anemones Metridium dianthus (Ellis) and Adamsia palliata (Bohad) ). Предположение, что это повторение (рекапитуляция) пройденного эволюционного пути, выглядит вполне правдоподобно.
В других группах кораллов (например, у восьмилучевых) личинки становятся лецитотрофными. Рта и кишечника у таких личинок нет. А у более эволюционно молодых стрекающих в жизненном цикле появляется стадия медузы. Личинки обладателей медуз — планулы — сильно упрощены, рта у них не бывает никогда. Это понятно: медуза сама прекрасно умеет расселяться, так что роль планктонной личинки в этом деле уменьшается и тратить ресурсы на создание сложных личинок больше не стоит.
Слишком сложная диплеврула
Билатерально-симметричные животные — сестринская группа стрекающих. Решающим «изобретением» стрекающих, благодаря которому они и получили свое название, стал книдоцит — стрекательная клетка с выбрасывающейся ядовитой нитью. Появление книдоцитов сделало стрекающих очень эффективными хищниками и, вероятно, в какой-то мере компенсировало им отказ от подвижности. У билатерий такого преимущества на старте не было, и они продолжили ползать по дну в поисках добычи, с которой смогли бы справиться.
Есть все основания полагать, что жизненный цикл древних билатерий был точно таким же, как и у древних стрекающих: пелаго-бентическим, с планктонной личинкой и донной взрослой стадией. Потребность к расселению у ползающих билатерий никуда не делась, а потому никуда не делась и плавающая личинка. Казалось бы, вот вам и объяснение наличия необычно выглядящих планктонных личинок у современных морских животных. Однако стоит присмотреться к этим личинкам чуть внимательнее, как становится ясно, что всё не так просто.
Все билатерально-симметричные животные (за исключением проблематичных ксенацеломорф; см. Ксенотурбеллиды оказались близки к предкам двусторонне-симметричных животных, «Элементы», 15.02.2016) делятся на два огромных эволюционных ствола: первичноротые и вторичноротые. И у тех и у других личинки бывают очень разными: каждый видел, например, бескрылую личинку кузнечика или водную личинку лягушки — головастика. Нас сейчас, однако, интересуют только «первичные» личинки, претендующие хоть на какую-то преемственность с пресловутой геккелевской гастреей. Многообразие таких личинок часто сводят к двум основным типам (конечно, имея в виду, что это лишь первое приближение; см., например: В. В. Малахов, 1996. Происхождение хордовых животных). Типичная планктонная личинка первичноротых называется трохофорой, а типичная планктонная личинка вторичноротых — диплеврулой. Образцовые трохофоры есть у некоторых кольчатых червей и моллюсков. Что касается диплеврулы, то это общее название ранних личинок иглокожих и полухордовых, которые потом превращаются в торнарий (у кишечнодышащих), бипиннарий (у морских звезд) и так далее.
У всех этих личинок есть признаки, которые никак не позволяют вывести их прямо из геккелевской гастреи (в отличие от личинок кораллов). Во-первых, гастрея радиально-симметрична, а у трохофор и диплеврул симметрия билатеральная — как у взрослых животных. Во-вторых, трохофоры и диплеврулы несут компактные группы специализированных клеток с мощными ресничками, которые называются ресничными шнурами. В общем, эти личинки явно прошли свой собственный непростой эволюционный путь.
Радиально-симметричными (и то лишь внешне) у билатерий бывают только самые ранние личинки — например, свободно плавающие бластулы, которые встречаются у морских животных. Их симметрия напоминает о древнейших временах, когда билатерий, что называется, еще и духу не было. Но ответа на вопрос о происхождении трохофор и диплеврул это не дает.
Воспари ввысь, стань личинкой
Академик Владимир Васильевич Малахов с группой коллег (из которых хочется особенно отметить Елену Николаевну Темереву) недавно опубликовал статью, в которой изложил свой сценарий происхождения планктонных личинок билатерий.
Откуда у трохофор и диплеврул билатеральная симметрия? Ответ очевиден: от взрослой формы, которая уже адаптировалась к ползанью по дну. Отсюда один шаг до предположения, что билатерально-симметричная личинка и есть юная (по-научному говоря — ювенильная) версия донной взрослой формы, в какой-то момент оторвавшаяся от дна и поднявшаяся в толщу воды.
Очень похожие события совершенно точно происходили в эволюции кораллов. У одиночных шестилучевых кораллов цериантарий поздняя личинка, продолжающая плавать в толще воды, представляет собой довольно продвинутое животное с таким числом щупалец и внутренних перегородок, каким в других группах кораллов обладают особи, уже прошедшие метаморфоз и живущие на дне. Очевидно, у цериантарий исходно донная ювенильная стадия поднялась со дна и превратилась в планктотрофную личинку. Удлинение личиночной части жизненного цикла за счет перехода донных ювенильных стадий в планктон — эволюционное явление, которое Малахов и его коллеги назвали термином «ларвализация».
Вполне может быть, что подобная ларвализация произошла и в эволюции билатерий (рис. 3). Во всяком случае, это сразу объясняет тип симметрии трохофор и диплеврул. Билатерально-симметричные личинки билатерий — это бывшие донные ювенильные формы, поднятые в толщу воды.

Рис. 3. Ларвализация у билатерий. В жизненном цикле А единственные плавающие стадии — это бластула и гаструла. В жизненном цикле Б бывшая донная ранняя ювенильная стадия поднялась в толщу воды (красная стрелка) и стала плавающей планктотрофной билатерально-симметричной личинкой (красная звездочка). Иллюстрация из обсуждаемой статьи в «Онтогенезе», с изменениями
Гораздо сложнее объяснить происхождение личиночных ресничных шнуров — структур, которые многие классические зоологи считали очень важными для понимания путей эволюции животного мира.
Версия Нильсена
В 1985 году датские зоологи Клаус Нильсен (Claus Nielsen) и Арне Нэрреванг (Arne Norrevang) опубликовали собственную гипотезу, объясняющую происхождение трохофор и диплеврул (C. Nielsen, A. Norrevang, 1985. The trochaea theory: an example of life cycle phylogeny). Попутно Нильсен, всегда обожавший планктонных личинок, построил на основе этой гипотезы целый детальный сценарий эволюции многоклеточных животных (C. Nielsen, 1985. Animal phylogeny in the light of the trochaea theory). Теорию Нильсена и Нэрреванга мало кто принял безоговорочно, но ее отдельные моменты убедили многих. И уж во всяком случае нельзя не признать, что она послужила хорошей основой для дальнейших обсуждений темы морских личинок и их эволюционного значения. Изложим ее коротко.
Начнем с того, что у всех без исключения многоклеточных животных когда-то были предки, которые несли реснички. Следует пояснить, что одиночные реснички называются жгутиками. Четкой границы между жгутиками и ресничками не существует (они устроены одинаково), но обычно о ресничках говорят, когда их много и они согласованно бьют в одну сторону. У одноклеточных предков животных жгутик был один. У губок он тоже один на каждой жгутиковой клетке (и у взрослых губок, и у личинок). Вероятно, и у геккелевской гастреи покровные клетки были классическими жгутиковыми. Однако в некоторый момент, который мы не можем пока датировать, но точно знаем, что он так или иначе был, — так вот, в некоторый момент вокруг первичного рта образовалось кольцо специализированных многоресничных клеток, задачей которых было гнать пищевые частицы в сторону кишки. Так возник первый ресничный шнур, который называется археотрохом.
Пока гастрея (уже не геккелевская, а более продвинутая) оставалась плавающей, археотрох был строго кольцевым. Но в эволюционной ветви, ведущей к билатериям, гастрея довольно скоро опустилась на дно и перешла к ползанью. Двигалась она аборальным (антиротовым) концом вперед, потому что на этом конце находился чувствительный центр. Ротовой конец, таким образом, оказался задним, что было для донного животного не слишком удобно. Поэтому рот у донного потомка гастреи начал вытягиваться на брюшную сторону. И окружающий его археотрох — тоже. В результате возник длинный щелевидный рот (переходная эволюционная стадия, до сих пор дающая себя знать в эмбриональном развитии многих беспозвоночных; см., например: Вторичноротость может оказаться первичной для двусторонне-симметричных животных, «Элементы», 06.09.2015), который в конце концов закрылся посредине, разделившись на два отверстия: ротовое впереди и анальное сзади.
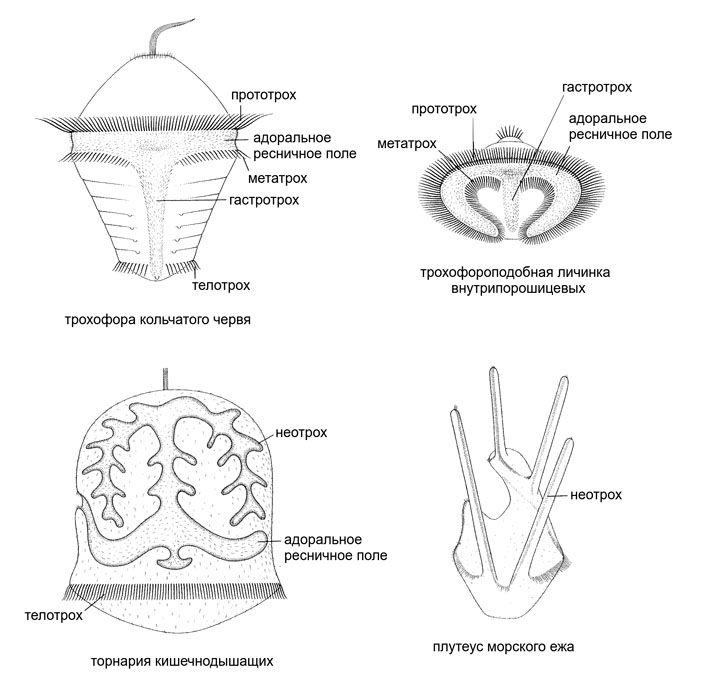
Что же при этом произошло с археотрохом? Он остался замкнутым контуром, окружающим со всех сторон бывший первичный рот. Но форма его, конечно, усложнилась. Петля археотроха, охватывающая сохранившийся рот (дефинитивный), вытянулась в поперечном направлении. Часть этой петли, находящаяся впереди от рта, в классической зоологической терминологии называется прототрохом, а часть, находящаяся позади от рта — метатрохом. Мощные реснички прототроха и метатроха бьют навстречу друг другу, загоняя пищевые частицы в рот. Прототрох служит заодно и для плавания, а метатрох задействован только в питании. Петля археотроха, охватывающая анальное отверстие, называется телотрохом. И наконец, там, где щелевидный первичный рот закрылся, от бывшего археотроха осталась соединяющая метатрох и телотрох срединная полоса ресничных клеток, которая называется гастротрохом. Мы получили типичную ресничную личинку первичноротых — трохофору (рис. 4; см. также рис. 1 и картинку дня: Трохофора, «Элементы», 04.10.2016).

Рис. 4. Типы личинок и соотношение осей тела по Нильсену — Нэрревангу. У первичноротых показаны трохофора и взрослое животное, у вторичноротых — диплеврула и взрослое животное, у стрекающих для сравнения медуза и полип. Всюду обозначена орально-аборальная ось бывшей гастреи: О — оральный конец (ротовой), А — аборальный конец (антиротовой). У стрекающих орально-аборальная ось переворачивается, когда полип становится медузой. У вторичноротых она остается на месте, хотя новый рот возникает вблизи аборального конца тела. А вот у первичноротых орально-аборальная ось сильно искривляется — это связано со смещением эмбрионального первичного рта (бластопора) на брюшную сторону. Стоит обратить внимание, что у диплеврулы помимо рта обозначены парные жаберные щели (см. далее). Остальные пояснения — в тексте. Иллюстрация из статьи: C. Nielsen, A. Norrevang, 1985. The trochaea theory: an example of life cycle phylogeny. См. также рис. 5 в статье: Современный анализ типов развития морских беспозвоночных подтверждает выдвинутую Геккелем теорию гастреи, «Элементы», 30.09.2013
Ресничные клетки важны для движения, но в то же время они легко преобразуются в чувствительные элементы (как в нашем ухе и глазу). Поэтому неудивительно, что с ресничными шнурами исторически связана закладка центральной нервной системы. Во всяком случае, Нильсен в этом убежден. В области прототроха и метатроха возникает окологлоточное нервное кольцо, а в области гастротроха — брюшная нервная цепочка.
Конечно, нильсеновская красивая схема воплощается в реальность по-разному. Гастротрох, телотрох и даже метатрох присутствуют у реальных трохофор далеко не всегда. С другой стороны, у трохофор иногда бывают добавочные ресничные пояски — паратрохи (между прототрохом и метатрохом) или мезотрохи (между метатрохом и телотрохом). Бывает и акротрох впереди от прототроха. Все это относится только к личинкам: у взрослых животных архаичная система ресничных шнуров вообще исчезает (за исключением коловраток, которых иногда считают неотеническими трохофорами, но с этой гипотезой согласны не все зоологи). Но главная проблема заключается в том, что изложенный сценарий подходит только к первичноротым. Почему?
Против течения
Дело в том, что личинки вторичноротых — диплеврулы — сильно отличаются от трохофор. Во-первых, клетки ресничных шнуров у них, как правило, не многоресничные, а жгутиковые (как мы знаем, это более примитивное состояние; см. C. Nielsen, 1987. Structure and function of metazoan ciliary bands and their phylogenetic significance). Во-вторых, ресничный шнур, окаймляющий рот, работает у диплеврулы по другому принципу. У трохофоры все реснички прототроха и метатроха бьют в сторону рта и, естественно, гонят туда воду вместе с микроскопическими пищевыми объектами. Это простая и очевидная система захвата пищи, которая называется downstream-системой (английское наречие downstream означает ‘вниз по течению’). Действительно, в downstream-системе пища просто плывет по течению, которое создают ресничные клетки. Но у диплеврулы клетки ресничного шнура, окружающего рот, бьют жгутиками в противоположную сторону — так, чтобы гнать воду от рта прочь. И только при обратном движении жгутиков они отбрасывают в сторону рта попадающиеся пищевые частицы. Это называется upstream-системой (‘вверх по течению’). Как видим, способы захвата пищи у трохофор и диплеврул отличаются настолько сильно, что для них трудно предположить общее происхождение.
Нильсен и Нэрреванг прекрасно понимали эту проблему. И они построили для личинок вторичноротых отдельный эволюционный сценарий, опираясь на два факта.
- У личинок кишечнодышащих (но не иглокожих) есть дополнительный замкнутый ресничный шнур, охватывающий кольцом анальное отверстие. Его функция — чисто двигательная, а клетки в нем многоресничные, мощные, специализированные. Эта структура очень похожа на телотрох трохофоры, и вот тут эволюционная связь как раз вероятна.
- Диплеврула вторичноротых по своему плану строения во многих отношениях похожа на гребневика (см., например: К. В. Беклемишев, 1979. Зоология беспозвоночных). Раньше это сходство считали свидетельством близкого родства, сейчас понятно, что оно скорее всего конвергентное, но в любом случае оно есть. А у гребневиков имеются поры, сквозь которые из пищеварительной системы выходит вода. Они находятся на аборальном (антиротовом) конце тела, в области, где кишка гребневика разделяется на четыре симметричные ветви (правда, порами открываются только две из них). Почему бы у предков вторичноротых не быть подобным порам, и почему бы им не послужить преадаптацией для чего-нибудь нового?
По Нильсену и Нэрревангу, предком вторичноротых была радиально-симметричная гастрея с первичным ртом, окруженным археотрохом, которая в какой-то момент опустилась на дно и стала ползать на боку, аборальным концом вперед. С этого момента эволюция пошла иначе, чем у первичноротых (рис. 5). Функцию рта приняла на себя одна из аборальных пор, аналогичных аборальным порам гребневиков — та, которой повезло оказаться на стороне, ставшей брюшной. Так возник вторичный рот. Две другие аборальные поры, оказавшиеся по бокам, вероятно, стали жаберными щелями (альтернативные мнения см. Жаберные мешки вторичноротых могли произойти от органов выделения, «Элементы», 14.09.2015). Задачу сдвига рта вперед вторичноротым просто не пришлось решать. Поэтому никакого вытягивания первичного рта на брюшную сторону у них не случилось. Вместо этого первичный рот целиком превратился в анальное отверстие, а окружавший его археотрох целиком стал телотрохом. А вот вокруг вторичного рта возник совершенно новый замкнутый ресничный шнур — неотрох. Он не связан по происхождению с археотрохом, потому и работает по-другому.

Рис. 5. Происхождение диплеврулы по Нильсену — Нэрревангу. Предполагается, что у радиально-симметричного предка вторичноротых было 4 гастральных поры, симметрично расположенных на аборальном конце тела. Когда этот предок лег на грунт, пора, оказавшаяся на стороне, обращенной к грунту (то есть на брюшной), стала ртом. Две поры, оказавшиеся на боках, стали жаберными порами, они же жаберные щели. А пора, оказавшаяся на спине, обособилась вместе с частью первичной кишки. Эта часть первичной кишки (бывшая) стала головной целомической полостью, а бывшая гастральная пора — ведущим в нее отверстием (гидропором). Гипотеза насчет гидропора — самая спорная часть предложенного Нильсеном и Нэрревангом эволюционного сценария, но и она подтверждается кое-какими эмбриологическими данными. Остальные пояснения — в тексте. Иллюстрация из статьи: C. Nielsen, A. Norrevang, 1985. The trochaea theory: an example of life cycle phylogeny
План строения диплеврулы всегда был очень важен для биологов, интересующихся путями эволюции животного царства. Иногда диплеврулу даже рассматривают как архетип вторичноротых (см., например: Л. В. Белоусов, 1980. Введение в общую эмбриологию). Нильсен, правда, считал, что общий предок вторичноротых во взрослом состоянии был наиболее похож не на диплеврулу, а на позднюю личинку крыложаберного Cephalodiscus. Эта личинка имеет прямой кишечник (в отличие от взрослых крыложаберных, у которых он изогнут в связи с сидячей жизнью в трубке), единственную пару очень просто устроенных жаберных щелей (в отличие от кишечнодышащих и хордовых, у которых жаберных щелей много и они превращены в сложный фильтрационный аппарат) и систему околоротовых ресничных щупалец, которые захватывают пищу по upstream-механизму и, вероятно, представляют собой разрастания неотроха. Ранняя личинка, правда, у крыложаберных упрощена. Но зато у кишечнодышащих сохранилась планктотрофная личинка с максимальным числом древних признаков (у этой группы она называется торнарией). Так из структурных элементов, сохранившихся на разных стадиях жизненного цикла в разных группах животных, можно, как из деталей конструктора, собрать более-менее правдоподобный образ общего предка крупной эволюционной ветви — вторичноротых в данном случае. А ведь в число вторичноротых входят и позвоночные. И без плана строения диплеврулы тут не обойтись. Именно поэтому так интересно, откуда она взялась.
Версия Малахова
Малахов и его коллеги, признавая заслуги Нильсена, тем не менее предлагают свою версию того, как возникла диплеврула. Они сравнивают пространственное положение ресничных структур у трохофор и диплеврул, стремясь выявить их общий план строения — если, конечно, такой есть. В конце концов, самым ходовым критерием гомологии (то есть сходства, вызванного общим происхождением) всегда был критерий положения: «Гомология следует при одинаковом положении в сравниваемых конструкционных системах» (Ю. В. Мамкаев, 2012. Гомология и аналогия как основополагающие понятия морфологии). Для удобства можно представить личинку в виде глобуса и сделать карту в меркаторской проекции (рис. 6).


Рис. 6. Сравнение ресничных личинок первичноротых и вторичноротых. Верхний ряд — пространственное расположение ресничных структур трохофоры и диплеврулы (пояснения в тексте). Гастротрох и невротрох — синонимы, АРП — адоральное ресничное поле. Нижний ряд — эволюция способов захвата пищи. На помещенных здесь схемах личинок жирные стрелки обозначают направление биения ресничек и тока воды, а тонкие стрелки — направление движения пищевых частиц по адоральному ресничному полю. Все личинки изображены с брюшной стороны. Иллюстрации из обсуждаемой статьи в «Онтогенезе», с изменениями
Начнем с трохофоры. Ресничное вооружение типичной трохофоры состоит из трех компонентов, образующих непрерывный контур вокруг бывшего первичного рта (который у зародышей и личинок называется бластопором). Во-первых, это адоральное ресничное поле, окружающее рот и простирающееся от него направо и налево. У планктотрофных личинок края адорального ресничного поля окаймлены мощными ресничными шнурами — это уже знакомые нам прототрох и метатрох. У непитающихся лецитотрофных личинок прототрох и метатрох, как правило, не выражены. Во-вторых, это ресничная полоска, идущая по брюшной стороне вдоль линии замыкания бластопора — гастротрох (он особенно широк у ползающих личинок). И в-третьих, это ресничное кольцо вокруг ануса — телотрох. Развертка всех этих ресничных образований на плоскость будет напоминать типографскую букву «Т», где верхняя перекладина — это разросшееся в поперечном направлении адоральное ресничное поле, вертикальная палочка — гастротрох, а коротенькая нижняя перекладина — телотрох.
Теперь рассмотрим личинку кишечнодышащих — торнарию. У нее тоже есть (1) адоральное ресничное поле, окаймленное ресничными шнурами (неотрохом в терминологии Нильсена), (2) ресничная полоска, проходящая по средней линии брюшной стороны — невротрох и (3) кольцевой телотрох вокруг ануса. Добавим, что невротрох — это синоним нильсеновского гастротроха (см. О. М. Иванова-Казас, 1977. Сравнительная эмбриология беспозвоночных животных: трохофорные, щупальцевые, щетинкочелюстные, погонофоры). Разумеется, реальная живая природа всегда будет разнообразнее любых схем, сколь угодно реалистичных. У планктотрофных торнарий, как и у личинок иглокожих, конфигурация адорального ресничного поля и неотроха может очень сильно усложняться (см. рис. 1). Невротроха и телотроха у личинок иглокожих нет. Но у классической торнарии (не слишком поздней) перечисленные ресничные структуры образуют в развертке почти такую же Т-образную конструкцию, как у классической трохофоры.
На этом основании Малахов делает вывод, что системы ресничных шнуров трохофоры и торнарии полностью гомологичны друг другу. И система ресничных шнуров трохофоры (прототрох + метатрох + гастротрох + телотрох), и система ресничных шнуров торнарии (неотрох + невротрох + телотрох) суть одна и та же преобразованная замкнутая ресничная структура, окружавшая вытянутый щелевидный первичный рот у древнего предка билатерий. Что это была за структура? По мнению Малахова, она, в свою очередь, гомологична внутреннему кругу щупалец коралловых полипов (см. В. В. Малахов, 2016. Симметрия и щупальцевый аппарат книдарий). То, что у вторичноротых клетки неотроха остаются жгутиковыми, легко объяснить большей примитивностью этих животных. Телотрох торнарии — исключение, вероятно потому, что его функция — двигать все тело, а это предполагает большую гидродинамическую нагрузку.
Гипотеза, обоснованная таким образом, выглядит стройно и имеет свои следствия. В частности, из нее должно следовать, что у вторичноротых вытянутый на брюшную сторону бластопор все-таки был (см. Высказана гипотеза о происхождении вторичноротых от первичноротых, «Элементы», 08.12.2015). Будем помнить, что одной из главных идей сравнительной анатомии всегда была «идея единства плана строения, основанная на установлении неочевидного сходства» (Ф. Я. Дзержинский, 2005. Сравнительная анатомия позвоночных животных). Вот это неочевидное сходство мы и видим на картах трохофоры и торнарии, нарисованных Малаховым в меркаторской проекции.
В то же время надо отдавать себе отчет, что «малаховский» эволюционный сценарий противоречит «нильсеновскому» (во всяком случае, в его изложенном здесь варианте — взгляды Нильсена потом менялись, но это другой вопрос). Верны одновременно они быть не могут.
Что касается downstream- и upstream-систем планктотрофного питания, то главный вопрос в том, какая из них эволюционно первична. А может быть, никакая? Мы ведь не знаем, как питались общие предки билатерий (а уж тем более — общие предки билатерий и стрекающих). Если они собирали пищевые частицы со дна, то реснички на обращенной к этому дну поверхности тела, скорее всего, были недифференцированы. А вот после того, как бывшая ювенильная стадия ларвализовалась, поднялась в толщу воды и перешла к планктотрофному питанию, — тогда-то и произошла специализация ресничного аппарата. Нет ничего удивительного в том, что у первичноротых и вторичноротых она произошла по-разному: в одном случае сформировался downstream-, а в другом upstream-механизм. Если (как считает Малахов) обе версии ресничных шнуров гомологичны внутреннему кругу щупалец стрекающих, то чисто функциональные различия не препятствуют их гомологии.
Итак, на первый взгляд может показаться, что у диплеврул и трохофор очень много общего: и те и другие — полупрозрачные морские личинки, билатерально-симметричные, с ресничными шнурами, ну что еще нужно? Более детальные исследования, в которых огромную роль сыграл все тот же Клаус Нильсен, показали, что на самом деле различия между личинками первичноротых и вторичноротых очень глубоки. Появилась тенденция предполагать их совершенно независимое происхождение. А теперь намечается своего рода «отрицание отрицания»: мы видим, что общее между двумя типами личинок все же есть, и не на уровне общих функций, а на уровне вполне конкретных планов строения. Подтвердится ли эта идея и, главное, как она отразится на представлениях об эволюции билатерий в целом, мы, скорее всего, в ближайшие годы увидим.
Источник: elementy.ru





