













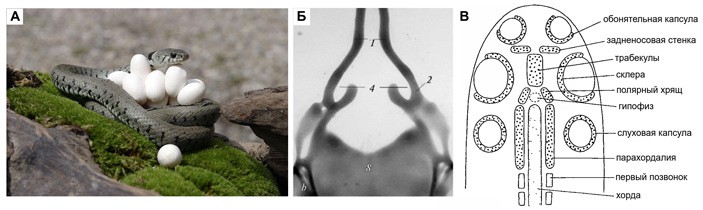


Рис. 1. А — самка обыкновенного ужа на кладке яиц; фото © M. Watson с сайта fineartamerica.com. Б — хрящевой череп эмбриона ужа (вид сверху) на стадии, когда образовались сонные вырезки (4). Здесь прекрасно видны трабекулы (1) и полярные хрящи (2). За сонными вырезками находится окно, позади которого — базальная пластинка (8), образованная слившимися задними частями парахордалий. Поперечной стойки тут еще нет. Иллюстрация из статьи H. V. Sheverdyukova, 2012. Polar cartilages and formation of crista sellaris in grass snake, Natrix natrix (Ophidia, Colubridae), chondrocranium at the early stages of embryogenesis. В — схема эмбриональных закладок нейрокраниума по Малькольму Джолли (Malcolm Jollie); рисунок из статьи M. T. Jollie, 1977. Segmentation of the vertebrate head. В отличие от обсуждаемой в тексте широко распространенной схемы Шмальгаузена, у Джолли между трабекулами и парахордалиями показаны полярные хрящи. Кроме того, трабекулы тут слиты в непарный элемент — так действительно бывает, хотя и далеко не у всех позвоночных (у змей они сливаются, но только впереди)
Детальные исследования индивидуального развития черепа обыкновенного ужа показали, что основная клиновидная кость (важная часть мозговой коробки) может у этой змеи развиваться разными способами. У большинства эмбрионов в образовании этой кости участвует непарный скелетный элемент, называемый поперечной стойкой или спинкой седла, но примерно в 15% случаев его вообще нет и основная клиновидная кость формируется иначе. Высказано предположение, что спинка седла — это архаичная структура, находящаяся прямо сейчас в процессе редукции.
…и гибкий змеиный хрящ, который хрустящ…
Дмитрий Быков
Сравнительная анатомия позвоночных — одна из самых заслуженных биологических наук. Она до сих пор полна загадок. Сейчас, в начале XXI века, многие проблемы сравнительной анатомии обретают новый смысл в результате взаимодействия с другими бурно развивающимися науками — биологией развития, молекулярной филогенетикой, палеонтологией. Но и классические методы по-прежнему позволяют узнать много интересного — если знать, что искать. И, конечно, одной из самых привлекательных частей организма позвоночного для сравнительных анатомов остается скелет головы, то есть череп.
Общий план
Начнем с того, что череп — это многокомпонентная структура. Мы привыкли считать, что внутри черепа находится головной мозг. Это, конечно, верно. Часть черепа, защищающая головной мозг, называется осевым черепом или нейрокраниумом. Кроме «коробки» вокруг мозга, в состав нейрокраниума обязательно входят парные капсулы органов чувств: слуховые капсулы, заключающие в себе перепончатые лабиринты внутреннего уха, и обонятельные капсулы, в которых находятся обонятельные мешки. Внутреннее ухо целиком замуровано внутри нейрокраниума, а обонятельные мешки, как легко догадаться, открываются наружу ноздрями.
Однако, помимо органов чувств, на голове находится рот. У позвоночных он ведет в глотку, пронизанную жаберными щелями — парными отверстиями, сквозь которые вода может вытекать наружу (рис. 2, А; см. также Жаберные мешки вторичноротых могли произойти от органов выделения, «Элементы», 14.09.2015). У водных позвоночных жаберные щели присутствуют всю жизнь, а у наземных они есть только на ранних стадиях, личиночных (амфибии) или эмбриональных (рептилии, птицы, млекопитающие). Глотка имеет собственный скелет, который называется висцеральным черепом или спланхнокраниумом. Часть висцерального черепа, находящаяся между двумя последовательными жаберными щелями, называется висцеральной дугой. Одна из висцеральных дуг — челюстная — у большинства позвоночных превращена в аппарат захвата пищи, то есть в челюсти. На других висцеральных дугах сидят органы водного дыхания — жабры. У рыб эти дуги сложно расчленены и подвижны, а у наземных позвоночных они редуцируются. Но и у человека, кроме нижней челюсти, сохранились еще три группы подвижных элементов спланхнокраниума: подъязычный аппарат, косточки среднего уха и гортань. Это и есть остатки жаберных дуг.

Рис. 2. А — общий план строения рыбы. Б — наиболее общепринятая схема эмбриональных закладок нейрокраниума (вид сверху). В — ископаемая кистеперая рыба эустеноптерон (Eusthenopteron). Г — схема нейрокраниума типичной кистеперой рыбы (вид сбоку). Стоит обратить внимание на толстую хорду, которая пронизывает задний блок мозговой коробки и упирается в передний. Д — нейрокраниум латимерии с подчерепной мышцей (вид сбоку). Е — отделы нейрокраниума по Шмальгаузену. Источники рисунков: А — статья A. S. Romer, 1972. The vertebrate as a dual animal — somatic and visceral; Б, Е — книга И. И. Шмальгаузен, 1947. Основы сравнительной анатомии позвоночных животных; В — блог boku-sui.blogspot.com; Г — книга Ф. Я. Дзержинский, 2005. Сравнительная анатомия позвоночных животных; Д — статья: H. Dutel et al., 2013. A reevaluation of the anatomy of the jaw-closing system in the extant coelacanth Latimeria chalumnae
Сейчас нас интересует в основном нейрокраниум. Его строение легко свести к стандартной схеме, показанной на рис. 2, Б (см., например: Ф. Я. Дзержинский, 2005. Сравнительная анатомия позвоночных животных). По бокам от хорды, которая у зародышей позвоночных не только тянется сквозь туловище, но и заходит в голову, возникают удлиненные хрящи — парахордалии. Впереди от хорды примерно в то же время появляется другая пара удлиненных хрящей — трабекулы. Срастаясь вместе, парахордалии и трабекулы образуют дно мозговой коробки, или основание черепа. Вверх от них устремляются потоки клеток, которые формируют стенки мозговой коробки, и в последнюю очередь образуется ее крыша. Кроме того, на стадии формирования стенок в мозговую коробку вливаются слуховые и обонятельные капсулы, которые становятся ее неотъемлемыми частями.
Область обонятельных капсул и глазниц часто называют трабекулярной частью осевого черепа, а область слуховых капсул и затылка — парахордальной частью. Интересно, что у кистеперых рыб (рис. 2, В), включая современную латимерию, эти части до сих пор соединены подвижно (рис. 2, Г). Положение трабекулярного блока относительно парахордального у них регулируется мощной подчерепной мышцей (рис. 2, Д). Но у большинства других позвоночных парахордальный и трабекулярный блоки во взрослом состоянии слиты. Маркером границы между ними служит гипофиз — важнейшая эндокринная железа, связанная с дном промежуточного мозга (рис. 2, Е). Сам гипофиз всегда находится внутри трабекулярного блока, но близко к его задней границе. Хорда никогда не заходит вперед дальше гипофиза, даже если она сохраняется всю жизнь, как у той же латимерии.
Шведский палеонтолог Эрик Ярвик (Anders Erik Vilhelm Jarvik) убедительно показал, что трабекула кистеперых рыб и наземных позвоночных — это на самом деле фрагмент челюстной дуги, совершивший переход из висцерального черепа в осевой. Это означает, что трабекулярный отдел нейрокраниума в большой степени состоит из структур, заимствованных у спланхнокраниума. Проще говоря, передняя половина мозговой коробки по крайней мере частично имеет висцеральное (то есть жаберное) происхождение.
Откуда же взялась ее остальная часть? Вот тут нам потребуется краткое отступление, касающееся истории науки.
Двухсотлетняя загадка
В 1807 году 28-летний экстраординарный профессор Йенского университета Лоренц Окен (Lorenz Oken) в первой же своей лекции изложил теорию, согласно которой череп состоит из нескольких сросшихся позвонков. В том же году текст лекции был издан отдельной книжкой под названием «О значении костей черепа» (L. Oken, 1807. Über die Bedeutung der Schädelknochen). Независимо от Окена к той же самой идее пришел знаменитый поэт и естествоиспытатель Иоганн Вольфганг Гёте. Он начал размышлять над позвоночной теорией черепа раньше, но опубликовал ее позже — только в 1823 году. Так что приоритет здесь по справедливости принадлежит Окену, который опубликовал свою теорию сразу.
Позвоночная теория черепа проста и наглядна. Взглянув на череп какого-нибудь млекопитающего, мы сразу видим впереди носовые кости, потом лобные, потом теменные, потом затылочные. Почему бы не предположить, что это слившиеся позвонки, вполне сравнимые с позвонками туловища? Именно так авторы позвоночной теории и рассуждали. «Головной мозг есть сильно развитый спинной мозг, так же как мозговая коробка есть объемистый спинной хребет», — писал Окен. По тем временам это выглядело убедительно, и в первой половине XIX века позвоночная теория черепа стала среди анатомов более-менее общепринятой.
Проблема была в том, что ученые никак не могли прийти к согласию по вопросу: сколько же, собственно, в голове позвонков? Более того, с накоплением материала ясность парадоксальным образом только убывала. В 1834 году швейцарский анатом и палеонтолог Луи Агассис (Jean Louis Rodolphe Agassiz) выступил вообще против позвоночной теории черепа, причем его главным аргументом — внимание! — было то, что хорда у зародышей позвоночных никогда не проникает в мозговую коробку на всю длину последней. Часть черепа, расположенную впереди от хорды, Агассис считал «совершенно чуждой позвоночной системе».
Окончательно разрушил позвоночную теорию черепа знаменитый Томас Гексли (Thomas Henry Huxley). Он показал, что носовые, лобные и другие кости, которые сторонники этой теории сравнивали с позвонками, формируются в эмбриональном развитии настолько иначе, что их гомология с позвоночником полностью исключена (T. H. Huxley, 1859. On the theory of the vertebrate skull). После этого позвоночная теория черепа перестала быть актуальной — во всяком случае, в ее классическом варианте.
Но главный вопрос, волновавший сторонников и противников этой теории, остается в силе до сих пор, несмотря на то, что с начала полемики прошло уже 200 лет (см., например: S. Kuratani, 2008. Is the vertebrate head segmented?—evolutionary and developmental considerations). В современной форме этот вопрос звучит так: есть ли в голове позвоночных метамерия, то есть сериальная повторяемость целых комплексов органов? То, что она есть в туловище, не вызывает никаких сомнений: об этом наглядно свидетельствуют позвонки, спинномозговые нервы, мышцы спины и живота и некоторые другие структуры. А вот есть ли в голове что-нибудь подобное?
Тут мы и возвращаемся к парахордалиям и трабекулам. Замечательная петербургская исследовательница Елена Дмитриевна Регель (1915–2003) показала, что в ходе развития черепа самых примитивных современных хвостатых амфибий — углозуба и лягушкозуба — парахордалии образуют три пары утолщений, напоминающих хрящевые дуги позвонков. Причем рядом с ними возникают три пары маленьких мышц, очень похожих на метамерные мышечные блоки туловища. Добавим, что парахордалии, подобно позвонкам и в отличие от большей части черепа, образуются из групп эмбриональных клеток, называемых склеротомами. Поэтому есть все основания считать разрастания парахордалий гомологами дуг позвонков, а соответствующие части головы — ничем иным, как тремя головными метамерами. Один из них связан с подъязычной висцеральной дугой и лицевым нервом, другой — с первой жаберной дугой и языкоглоточным нервом, и последний — со второй жаберной дугой и блуждающим нервом. На этом первичная голова позвоночного, по-видимому, кончается (см. С. А. Ястребов, 2018. Метамерия головы позвоночных: современное состояние проблемы).
Итак, в отношении парахордальной части нейрокраниума позвоночная теория черепа, как ни удивительно, по существу верна. Однако данные по висцеральному скелету, черепно-мозговым нервам и целомическим образованиям указывают на то, что впереди от парахордальной части есть еще два метамера — челюстной и предчелюстной. Они тоже связаны с соответствующими висцеральными дугами (предчелюстная висцеральная дуга у позвоночных неполна, но ее существование достаточно надежно подтверждается и палеонтологией, и эмбриологией). Особенно же интересна область границы между двумя отделами осевого черепа, маркером которой, как мы помним, служит гипофиз. Какие скелетные структуры мы можем увидеть рядом с ним?
В окрестностях гипофиза
Скелет гипофизарной области — не такая уж terra incognita для сравнительных анатомов. Они интересуются этой темой еще со времен Мартина Генриха Ратке (Martin Heinrich Rathke), который прославился, открыв жаберные щели у зародышей млекопитающих и птиц. Ратке сделал и другие открытия: например, именно он впервые описал трабекулы, а вырост глотки, из которого образуется передняя доля гипофиза, до сих пор называется в его честь карманом Ратке. Казалось бы, при такой длинной истории исследований тут давно должно быть все ясно. Но нет, в гипофизарной области черепа до сих пор таятся проблемы, о которых авторы учебников (даже очень хороших) предпочитают умалчивать, чтобы не нарушать стройность рассказа.
В начале XX века несколько исследователей обнаружили, что у зародышей самых разных рыб — акул, панцирных щук, осетров — дно мозговой коробки в гипофизарной области образуют отдельные продолговатые хрящи, не являющиеся ни трабекулами, ни парахордалиями, но расположенные как раз между ними (рис. 1, В). Известный голландский сравнительный анатом Ян Виллем ван Вай (Jan Willem van Wijhe) назвал эти элементы полярными хрящами. В дальнейшем выяснилось, что полярные хрящи есть у рептилий и у птиц. Гипофизарные хрящи млекопитающих, возможно, тоже гомологичны полярным хрящам — во всяком случае, в анатомии человека это синонимы (R. F. Gasser, 1976. Early formation of the basicranium in man). Такая высокая эволюционная устойчивость означает, что перед нами важные элементы плана строения. Но откуда они взялись — было непонятно. Забегая вперед, можно сказать, что общепринятого ответа на этот вопрос нет и сейчас. Если происхождение трабекул и парахордалий в первом приближении известно, то полярные хрящи остаются загадкой.
Некоторую ясность в эту тему внесла серия великолепных работ по развитию черепа обыкновенного ужа (Natrix natrix, рис. 1, А), которые сделала сотрудница киевского Института зоологии имени Шмальгаузена Анна Владимировна Шевердюкова (Ганна Володимирівна Шевердюкова). Она описала развитие всего черепа ужа на всех стадиях, от первых перепончатых закладок до полного набора костей. Сейчас нас интересует только маленький фрагмент этой истории развития. Итак, помимо трабекул (впереди) и парахордалий (сзади), в состав хрящевого скелета гипофизарной области зародыша змеи входят еще два анатомических «действующих лица».
Во-первых, это уже знакомые нам полярные хрящи, расположенные между трабекулами и парахордалиями. Шевердюкова показала, что у змей они закладываются совершенно самостоятельно — раньше это не было точно известно.
Во-вторых, это поперечная хрящевая перекладина, которая отделяет гипофиз (впереди) от переднего конца хорды (сзади). Она располагается на уровне передних концов полярных хрящей, близ границы с трабекулами. Этот элемент скелета описан у множества позвоночных, от акул до человека, но называется он у разных авторов по-разному: постпитуитарная комиссура, или предушной мостик, или седловидный гребень, или спинка седла, или акрохордальный хрящ. Все это, по-видимому, синонимы. Перед нами еще одна структура, история изучения которой длинна, почтенна и загадочна. У человека и полярные хрящи, и спинка седла в конце концов входят в состав клиновидной кости. Углубление для гипофиза, ограниченное сзади спинкой седла (dorsum sellae), в анатомии человека традиционно называется турецким седлом (sella turcica).
Полярные хрящи у змеи, как уже сказано, закладываются самостоятельно. Но довольно скоро они сливаются и с парахордалиями, и с трабекулами, образуя вместе с ними единые длинные «продольные балки» дна мозговой коробки.
Кроме того, как раз между полярными хрящами (в том месте, где они контактируют с трабекулами) внутрь мозговой коробки зародыша змеи входят внутренние сонные артерии, снабжающие кровью головной мозг. Полярный хрящ сначала охватывает сонную артерию своим выростом, образуя сонную вырезку (рис. 1, Б), а потом эта вырезка замыкается и возникает хрящевое сонное кольцо.
После этого между сонными кольцами образуется поперечная хрящевая перемычка. Она располагается точно так же, как описанный у других позвоночных акрохордальный хрящ, и вполне могла бы быть его гомологом.
Но вот тут-то появляется трудность. Для ряда позвоночных, в том числе для черепах и для птиц, достаточно надежно установлено, что акрохордальный хрящ у них формируется как самостоятельный элемент, имеющий собственный центр охрящевения. Недавно то же самое удалось показать для прыткой ящерицы (Lacerta agilis; см. А. Н. Ярыгин, 2009. Закладка парахордалий, акрохордального хряща и черепных трабекул в эмбриогенезе прыткой ящерицы, Lacerta agilis (Reptilia, Squamata)). Проще говоря, акрохордальный хрящ — это не производное какого-то другого хряща, а отдельная структура с отдельной эволюционной судьбой. Еще один элемент, образующий дно осевого черепа, в придачу к парахордалиям, трабекулам и полярным хрящам.
Проблема в том, что у обыкновенного ужа поперечная хрящевая перемычка выглядит скорее как продукт встречных разрастаний полярных хрящей, чем как что-то самостоятельное (H. V. Sheverdyukova, 2012. Polar cartilages and formation of crista sellaris in grass snake, Natrix natrix (Ophidia, Colubridae), chondrocranium at the early stages of embryogenesis). Поэтому Шевердюкова предпочитает называть этот элемент не акрохордальным хрящом, а поперечной стойкой (transverse bar), — предельно нейтральным термином, не предполагающим никакой гомологии. В статье, опубликованной в 2012 году, она высказала мнение, что поперечная стойка ужа — это не гомолог, а аналог акрохордального хряща черепах, птиц и ящериц. Независимо возникшая структура, занимающая то же положение.
Может ли такое быть? В принципе вполне может. Однако, прежде чем разбираться дальше, давайте зададимся очевидным вопросом: почему это вообще заслуживает внимания? Могут ли хрящи черепа нерожденной змеи сообщить нам что-то важное об устройстве Вселенной?
Рыбы, ученые и архетипы
Начнем с самого начала. Все мы знаем, что живая природа очень разнообразна, причем сильнее всего ее разнообразие проявляется в бесчисленных формах организмов — особенно эукариотных и многоклеточных. «Если природа в отношении функциональных схем проявляет большую экономию, то в том, что касается форм живых существ и их органов, она безудержно расточительна и в этом уподобляется художнику, ни одно произведение которого не повторяет другое» (Б. С. Кузин, 1992. Упадок систематики). Биология упорядочивает это грандиозное разнообразие по двум направлениям: эволюционное древо она делит на таксоны, а каждый организм — на мероны (С. В. Мейен, Ю. А. Шрейдер, 1976. Методологические аспекты теории классификации). Простейший пример мерона — любой орган многоклеточного тела. Множество меронов — это множество частей организма, рассматриваемых как самостоятельные (хотя и подчиненные) объекты исследования. Морфолог, то есть исследователь биологической формы и ее превращений, всегда начинает с разбиения целого на отдельные мероны: иначе просто невозможен никакой научный анализ. Естественно, что иногда мероны сливаются (если под разными названиями была описана одна и та же структура) или дробятся (если структура, которую раньше считали элементарной, оказалась состоящей из нескольких элементов разного происхождения).
Все было бы просто, если бы таксономия и мерономия были альтернативными и независимыми способами познания. Но на самом деле они сильнейшим образом взаимно переплетены. Любой мерон, будь то жгутик эвглены или локтевая кость позвоночного, свойствен не всем живым организмам на Земле, а некоему таксону, границы которого должны быть четко очерчены. Мерономия по самой своей природе не бывает универсальной — она действует только внутри «своего» таксона. Совокупность меронов, инвариантная для данного таксона, называется архетипом этого таксона (С. В. Мейен, 1977. Таксономия и мерономия). Например, мы можем говорить об архетипе насекомых, архетипе моллюсков или архетипе позвоночных. Архетип — это локальный закон природы, область действия которого ограничена одним таксоном (С. В. Мейен, 1978. Основные аспекты типологии организмов).
Правильно составленный архетип обладает огромной предсказательной силой. Его перестройка может серьезно изменить представления о путях, которыми шла макроэволюция (как это произошло, когда Эрик Ярвик, основываясь на большом палеонтологическом материале, пересмотрел гомологии трабекул и некоторых других составных частей черепа рыб). И самый высокий риск таких изменений возникает как раз тогда, когда в составе сложной структуры обнаруживаются «лишние» элементы, статус которых неясен.
Скажем, чему гомологичны полярные хрящи? Морфологи начали биться над этой загадкой почти сто лет назад (E. P. Allis, 1923. Are the polar and trabecular cartilages of vertebrate embryos the pharyngeal elements of the mandibular and premandibular arches?), но высказанные тогда идеи были поставлены под сомнение новыми палеонтологическими открытиями. Шведский палеонтолог Ханс Бьерринг (Hans Christian Bjerring) предложил очень смелую гипотезу. Он гомологизировал полярный хрящ с подчерепной мышцей кистеперых рыб — той самой, которая связывает передний и задний блоки осевого черепа.
Дело в том, что по положению в черепе подчерепная мышца и полярный хрящ очень хорошо соответствуют друг другу. По мнению Бьерринга, после того, как подчерепная мышца стала не нужна, ее эмбриональный зачаток переключился на другое направление дифференцировки и превратился в скелетный элемент. У ископаемой кистеперой рыбы Youngolepis, у которой подвижность внутри мозговой коробки отсутствует, нет и регулирующей эту подвижность подчерепной мышцы. Но зато есть костный мостик, расположенный точно так же, как располагалась бы подчерепная мышца, если бы она была (H. C. Bjerring, 1985. Facts and thoughts on piscine phylogeny). Вероятно, это не что иное, как окостеневший полярный хрящ.
Сама подчерепная мышца, по Бьеррингу, представляет собой древний мышечный блок челюстного метамера. Проблема этой гипотезы в том, что у современной кистеперой рыбы — латимерии — подчерепная мышца очень длинная, она тянется от самого затылка (рис. 2, Д). Бьеррингу пришлось предположить, что у этой рыбы она стала составной — включила в себя мышечные блоки других головных метамеров (те самые, которые наблюдала Регель у личинок хвостатых амфибий; H. C. Bjerring, 1993. Yet another interpretation of the coelacanthiform basicranial muscle and its innervation). Недавние тщательные анатомические исследования латимерии неожиданно вполне подтвердили эту идею (см. А. Н. Кузнецов, 2018. Пояснения к статье Ф. Я Дзержинского «Загадка двухблочного черепа представителей Sarcopterygii. Ловушка для функциональных морфологов»). В конце концов, удивляться тут нечему: латимерия прошла долгий путь собственной эволюции (см. Можно ли считать латимерию живым ископаемым?, «Элементы», 01.03.2013). А у девонских кистеперых рыб подчерепная мышца гораздо более короткая, так что предположение о ее принадлежности к челюстному метамеру выглядит резонно.
Однако, пока полярный хрящ еще был мышцей, между трабекулами и парахордалиями должна была находиться какая-то другая скелетная структура. Очевидно, это был акрохордальный элемент, возникший из склеротома челюстного метамера подобно тому, как из склеротомов следующих метамеров возникают парахордалии и дуги позвонков (H. C. Bjerring, 1978. Contribution to structural analysis of the head of craniate animals). Вот вам и позвоночная теория черепа в современном облике. Когда появился полярный хрящ, он в какой-то мере потеснил и заменил более древний акрохордальный.
А теперь обратимся к тому, что может нам рассказать об этом змеиный череп.
Моментальное фото эволюции
Недавно вышла новая работа Анны Шевердюковой, написанная в соавторстве с ее учителем Михаилом Фотиевичем Ковтуном (Михайло Фотійович Ковтун). Ученые опять обратились к черепу ужа. Но теперь их заинтересовал не стандартный ход развития черепа, а его изменчивость.
Дело в том, что со скелетом гипофизарной области ужа все оказалось не так-то просто. Мы знаем, что хрящевая поперечная стойка там выглядит как парная структура, образуемая встречными разрастаниями полярных хрящей. Но перейдя к изучению более поздних стадий, когда в черепе появляются кости, Шевердюкова обнаружила, что независимый непарный центр окостенения в поперечной стойке все же есть (H. V. Sheverdyukova, 2017. Development of the osteocranium in Natrix natrix (Serpentes, Colubridae) embryogenesis I: development of cranial base and cranial vault). Таким образом, возникающая здесь кость — у рептилий она называется основной клиновидной — явно имеет очень сложное происхождение.
В новой работе Шевердюкова и Ковтун показали, что основная клиновидная кость ужа может развиваться как минимум двумя способами: (1) с окостенением поперечной стойки или же (2) только из встречных разрастаний полярных хрящей (рис. 3). Судя по всему, примерно у 15% ужиных эмбрионов поперечная стойка как самостоятельный элемент вообще не образуется. Это не задержка, а именно полное отсутствие. На строение основной клиновидной кости, возникающей в результате, выбор того или иного пути развития не влияет. Оба варианта могут наблюдаться не только внутри одной популяции, но и внутри одной яйцевой кладки, у братьев или сестер.

Рис. 3. Два альтернативных способа развития основной клиновидной кости ужа. Слева — два эмбриона на одной и той же стадии, где все элементы хрящевые. В одном случае (A) поперечная стойка сформирована, в другом случае (A’) ее нет. Справа — два эмбриона на более поздней стадии (опять одной и той же!), когда череп окостеневает: в одном случае (B) образуется самостоятельный акрохордальный центр окостенения, а в другом случае (B’) кость формируется исключительно за счет окостенений встречных отростков полярных хрящей. Обозначения: tc — трабекула, pc — полярный хрящ, cf — сонное кольцо, cs — спинка седла (= поперечная стойка), pf — гипофизарное окно, bf — базикраниальное окно (между парахордалиями), Ac — слуховая капсула, ob — центры окостенения, 1 — размытые границы хряща, который начал разрушаться и замещаться костью, 2 — формирующаяся надкостница. На фото B спинка седла значительно опережает по степени окостенения полярные хрящи, а на фото B’ она так и не образовалась. Длина масштабной линейки везде 0,5 мм. Иллюстрации из обсуждаемой статьи в Journal of Morphology
Такая сильная морфологическая вариабельность развития осевого черепа обнаружена у змей впервые. Вариант, когда независимая поперечная стойка все-таки есть, количественно преобладает, поэтому его можно условно считать «нормой». Но и отсутствие поперечной стойки нельзя считать патологией: оно не приводит абсолютно ни к каким нарушениям, основная клиновидная кость у таких особей получается точно такой же, как и в «нормальном» варианте. Это хороший пример давно известного в эмбриологии принципа эквифинальности: развитие может разными путями приводить к одному и тому же конечному результату (см., например: Л. В. Белоусов, 1980. Введение в общую эмбриологию).
Поперечная стойка змеи совпадает по положению с акрохордальным элементом (он же спинка седла и прочее). Вполне возможно, что это он и есть. Тут нет ничего удивительного, акрохордальный элемент типичен для черепа рептилий — крокодилов, черепах, ящериц. Причем у многих из них он достоверно развивается как независимая непарная структура. Тоже ничего удивительного, акрохордальному элементу так и положено.
По всей видимости, наличие самостоятельного и непарного акрохордального элемента является примитивным для рептилий признаком (O. Yaryhin, I. Werneburg, 2017. Chondrification and character identification in the skull exemplified for the basicranial anatomy of early squamate embryos). Тогда выглядит вполне логичным предположение, что у змей мы видим остаток этого элемента, находящегося прямо сейчас в процессе редукции. А поскольку этот процесс не завершен, то мы и наблюдаем во внутривидовой изменчивости ужа целый спектр состояний — от промежуточной стадии редукции до полного исчезновения. Исследование киевских ученых «поймало» план строения черепа змей непосредственно в процессе становления, in statu nascendi. Перед нами своего рода фотография, запечатлевшая событие, которое эволюционным морфологам очень редко удается увидеть. Но именно из подобных микрособытий и складывается макроэволюция — вплоть до таких масштабных явлений, как появление новых типов животных, полета или разума.
Добавим, что Шевердюкова и Ковтун подходят к проблеме очень аккуратно. Они в принципе не исключают, что непарная закладка поперечной стойки может оказаться новым приобретением змей, негомологичным классическому акрохордальному хрящу. Но все же они отдают предпочтение гипотезе редукции — видимо, обоснованно, учитывая, что редукция глазничного отдела нейрокраниума для змей вообще очень характерна.
Возвращаясь к разговору об архетипе, мы можем констатировать, что в гипофизарной области змеиного черепа теперь (если предыдущие рассуждения верны) найден полный набор его элементов: полярные хрящи и акрохордальный хрящ. Последний, правда, находится на пути к исчезновению. Увы, «река времен в своем стремленье» может унести что угодно, в том числе и важные элементы архетипа. Ни от изменчивости, ни от естественного отбора они ничем не защищены. Эволюция — это всегда процесс, в котором что-то возникает, а что-то исчезает бесследно.
Новая и древняя голова
Великий знаток сравнительной анатомии и эволюционной теории академик Иван Иванович Шмальгаузен, именем которого называется киевский Институт зоологии, писал, что парахордальную часть черепа (в которую заходит хорда) можно иначе назвать хордальным отделом, а трабекулярную часть черепа (в которую хорда не заходит) можно иначе назвать прехордальным отделом (рис. 2, Е). Это то самое деление, на которое за сто лет до Шмальгаузена обратил внимание Луи Агассис. Границей между хордальным и прехордальным отделами головы Шмальгаузен считал уже знакомый нам гипофиз. Он обратил внимание на различия в эмбриональном развитии этих отделов и на то, что в нервных структурах прехордального отдела нет никаких двигательных выходов. Иными словами, все эти структуры — сенсорные. Самые передние двигательные нервы выходят из среднего мозга, который расположен позади от гипофиза. Вывод: «Прехордальная часть головы представляет новообразование, развившееся в связи с развитием переднего мозга и соответствующих органов чувств» (И. И. Шмальгаузен, 1947. Основы сравнительной анатомии позвоночных животных).
Проницательностью Шмальгаузена можно восхититься. Прехордальная часть головы позвоночных до такой степени изобилует эволюционными новообразованиями, что известный американский нейробиолог Глен Норткат (Glenn Northcutt) предложил на этой основе популярную в наши дни «гипотезу новой головы» (G. R. Northcutt, 2005. The new head hypothesis revisited). Многие из этих новшеств сейчас изучаются на молекулярном уровне: например, эволюция переднего мозга, по-видимому, тесно связана с эволюцией регуляторных генов семейства Anf, которое появилось только у позвоночных (A. V. Bayramov et al., 2016. The presence of Anf/Hesx1 homeobox gene in lampreys suggests that it could play an important role in emergence of telencephalon).
Однако план строения позвоночных не возник из ничего. Позвоночные, как и все хордовые, являются потомками каких-то более древних вторичноротых. Как известно, кроме них к этой большой группе относятся иглокожие и полухордовые. Архетипом вторичноротых может считаться диплеврула — ранняя планктонная личинка иглокожих и полухордовых (см. Общее происхождение трохофор и диплеврул: за и против, «Элементы», 06.02.2020). Типичная диплеврула имеет три пары целомических мешков: протоцель (впереди рта), мезоцель (примерно на уровне рта) и метацель (в туловище). Тело всех нехордовых вторичноротых построено по этому плану: и у полухордовых, и у иглокожих есть протоцель, мезоцель и метацель (рис. 4, А). Для краткости можно называть соответствующие отделы тела протосомой, мезосомой и метасомой (см., например: J. Godeaux, 1974. Introduction to the morphology, phylogenesis and systematics of lower Deuterostomia).

Рис. 4. А — личинка полухордового накануне метаморфоза (вид сбоку). Строго говоря, это не диплеврула, а гораздо более поздняя личинка — торнария, но на роль «живого архетипа» подходит и она. Мезоцель и метацель у нее парные и расположены по бокам от кишки. Б — схема экспрессии Hox-генов в висцеральных дугах позвоночного. Изображены последовательные эмбриональные стадии. Чем интенсивнее цвет, тем больше Hox-генов экспрессируется в данной дуге. Предчелюстная и челюстная дуги белые: это область «передней головы», где экспрессии Hox-генов нет. В — схема экспрессии Hox-генов у молодого полухордового. В протосоме и мезосоме ее нет. Г — рисунок продольного разреза черепа латимерии, который использовала Е. Д. Регель, чтобы показать, что трабекулярный отдел черепа не тождествен прехордальному. Голубым цветом отмечены границы мезосомы согласно системе гомологий, предложенной П. П. Ивановым и Х. Бьеррингом. Источники рисунков: А — книга И. И. Шмальгаузен, 1947. Основы сравнительной анатомии позвоночных животных; Б — статья S. Kuratani, 2005. Developmental studies of the lamprey and hierarchical evolutionary steps towards the acquisition of the jaw; В — статья J. Aronowicz, C. J. Lowe, 2006. Hox gene expression in the hemichordate Saccoglossus kowalevskii and the evolution of deuterostome nervous systems; Г — книга J. Millot, J. Anthony, 1958. Anatomie de Latimeria chalumnae
В первой половине XX века петербургский эмбриолог Пётр Павлович Иванов (1878–1942) обосновал идею, что план строения диплеврулы сохранился и у хордовых. В теле позвоночных протосоме диплеврулы соответствует предчелюстной метамер, мезосоме — челюстной метамер, а метасоме — все остальные метамеры, сколько бы их ни было. В начале XXI века это было подтверждено исследованиями генов семейства Hox, которые контролируют у животных отличия между сегментами (см. Программы работы Hox-генов у личинок и взрослых особей кольчатого червя принципиально отличаются, «Элементы», 27.05.2013). У позвоночных Hox-гены экспрессируются в области всех висцеральных дуг, кроме челюстной и предчелюстной (рис. 4, Б; см. S. Kuratani, 2005. Developmental studies of the lamprey and hierarchical evolutionary steps towards the acquisition of the jaw). А у полухордовых Hox-гены экспрессируются только в метасоме, причем схема их экспрессии там весьма похожа на то, что наблюдается у позвоночных (рис. 4, В, J. Aronowicz, C. J. Lowe, 2006. Hox gene expression in the hemichordate Saccoglossus kowalevskii and the evolution of deuterostome nervous systems). В протосоме и мезосоме полухордовых экспрессии Hox-генов нет.
Часть головы, не входящую в зону экспрессии Hox-генов, в современной биологической литературе называют уже устоявшимся термином «передняя голова» («anterior head»). Тут мы сталкиваемся с парадоксом. «Передняя голова» в самом деле богата эволюционными новшествами, но одновременно это очень архаичный отдел тела, некоторые особенности которого древнее самого типа хордовых. Ему-то, вероятно, и соответствует трабекулярный отдел черепа кистеперых рыб. Называть этот отдел «прехордальным» не стоило бы: в состав трабекулярного блока входит предполагаемый челюстной метамер, в область которого — иными словами, в область акрохордальных и полярных хрящей — хорда вполне себе заходит (рис. 4, Г). Гипофиз же действительно всегда расположен впереди от конца хорды. Хорошо известно, что в нервной трубке экспрессия Hox-генов заканчивается на границе между продолговатым и средним мозгом, а не между средним и промежуточным, как было бы, если бы рубеж «передней головы» проходил по гипофизу.
Гипофизарная область вообще является настоящим средоточием морфологических границ (R. H. Khonsari et al., 2013. The buccohypophyseal canal is an ancestral vertebrate trait maintained by modulation in sonic hedgehog signaling). Разобраться в этом важно не только ради предметного интереса к анатомии черепа, но и для лучшего понимания глубоких закономерностей эволюции планов строения многоклеточных животных. Здесь пригодятся и палеонтология, и генетика развития, и классическая морфология. И, конечно, хорошие исследования на эту тему надо всячески приветствовать.
Источник: elementy.ru




")
